Genetic and environmental causes of species range limits
Why species are limited in their geographic distribution is a central question in ecology evolution. During my PhD, I assessed recent theories stating that range edge population could suffer from the accumulation of deleterious mutation (mutational load) due to increased genetic drift, resulting from past range expansion or long-term isolation. I studied the dynamics of mutational load over the whole range of the North American plant species Arabidopsis lyrata in a species wide transplant experiment. I found that the expression of mutational load increased towards range limits in population with a long history of range expansion, and was further increased by a switch to selfing. These patterns suggest that further colonization or isolation at range limits may be impeded by further accumulation of mutational load, reducing performance below critical thresholds necessary to maintain persisting populations. Perrier et al., 2020, Evolution
In a follow up study, I assessed the environmental dependency of the expression of mutational load, to test whether range-edge population exposed to high drift would experience an increase in expression of mutational load when colonizing unsuitable habitats beyond range limits (Perrier et al., 2022, JEB) I also collaborated in two parallel studies in the framework of this research project, led by my colleague Dr. Darío Sánchez-Castro. The first study generally focused on testing whether the range limits of this species correspond to its niche limits (coming soon ...). The second tested the effect of genetic drift on local adaptation and environmental tolerance, with a focus on range limits (Sánchez-Castro et al., 2022, GEB). Further studies performed in the framework of this project included testing for pollination variation across this species range (Sánchez-Castro et al., Oecologia, in press). |

Arabidopsis lyrata subsp. lyrata in the wild
|

Wild population of Arabidopsis lyrata
(Picture by Darío Sánchez-Castro) |

Preparation of transplant experiment in five common garden experiments with colleague Darío Sánchez-Castro.
|
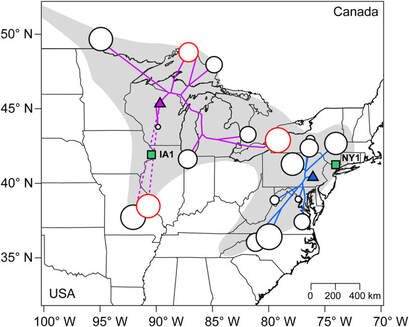
Mutational load expression (dot size) increases in edge population of each cluster (black line), with higher distance (purple and blue line) from core (triangles), and in selfing populations (red). Perrier et al., 2020, Evolution
|
Adaptation and speciationDuring my PhD, I also assessed what contributes to successful range expansion. I used Arabidopsis arenicola, the selfing sister species of A. lyrata, as study system. Despite sharing a common post-glacial ancestor with A. lyrata, A. arenicola colonized large subarctic and arctic regions of North America, providing a striking counter-example to the limitations on range expansion outlined above. I raised populations of both species and their laboratory generated hybrids in a climate chamber experiment simulating stress linked to growth in cold climates over a whole life cycle. To understand their divergent distribution, I tested whether A. lyrata and A. arenicola differ in their adaptation to cold temperatures and frost events (coming soon ...). In addition to their parapatric distribution, I further assessed if both species are isolated by other reproductive barrier contributing to adaptive differentiation I therefore tested the levels of intrinsic reproductive isolation by comparing performance of interspecific hybrids and their parental populations (Perrier and Willi, 2022, JEB).
|

Arabidopsis arenicola
|



Genetic diversity and systematics

|
Dianthus superbus
I collaborated in the revision of the four contested subspecies of the carnation Dianthus superbus, using morphometric, genotypic and ecological data. These subspecies actually form two distinct species, and further differentiation was linked to ecotype formation and plasticity. Hardion et al., 2020, Systematics and Biodiversity |

|
Miscanthus x giganteus
Using phenotypic and genotypic data, I showed that the genetic homogeneity of Miscanthus x giganteus crop fields in France is compromised by the co-occurrence of its invasive relative M. sacchariflorus, likely due to misidentification of both nursery and horticulture accessions. Perrier et al., 2019, Weed research |
Wetland ecology
|

|
Ecological restauration in the upper Rhine floodplain
During my Master thesis, I assessed the success of ecological restoration operations in a complex wetland ecosystem of the upper Rhine floodplain. By compiling survey and cartography data over 20 years, I showed that these operation improved the biodiversity, ecosystem functionality and ecosystem services. |
